
 French
French Deutsch
DeutschParabasalia , la enciclopedia libre
| Parabasalia | ||
|---|---|---|
 | ||
| Taxonomía | ||
| Dominio: | Eukarya | |
| Reino: | Protista | |
| (sin rango) | Excavata | |
| Filo: | Metamonada | |
| Clase: | Parabasalia Honigberg 1973 | |
| Órdenes[1] | ||
Parabasalia es una clase de protistas flagelados, la mayoría de los cuales son endosimbiontes, comensales o parásitos de animales.[2] Incluye una gran variedad de especies encontradas en el tubo digestivo de termitas y cucarachas, muchas de las cuales tienen bacterias simbióticas que les ayudan a digerir la madera. Otras especies son parásitas, incluyendo algunos patógenos para los seres humanos, como Trichomonas, Pentatrichomonas hominis y Dientamoeba fragilis. Parabasalia comprende unas 470 especies conocidas.[3]
Características[editar]
La mayoría de los parabasálidos son endosimbiontes, comensales o parásitos de invertebrados o vertebrados, y por lo general habitan en la parte distal del tubo digestivo de estos animales. Como excepción, se conocen algunas especies de vida libre. Muchas especies son simbióticas en el intestino de termitas y cucarachas, desempeñando un papel central en la digestión de la celulosa por parte de estos organismos. Por tanto, juegan un importante papel ecológico en la descomposición de los residuos vegetales en los ecosistemas terrestres y probablemente la relación simbiótica haya sido un elemento clave en la evolución del comportamiento social de sus anfitriones.[4] Otros parabasálidos son patógenos de considerable importancia médica y veterinaria. Con pocas excepciones, los parabasálidos no forman quistes.
Los parabasálidos son anaerobios y carentes de mitocondrias.[5] Se considera que esto es el resultado de una pérdida secundaria pues las células contienen pequeños hidrogenosomas que aparentemente se han desarrollado a partir de las mitoncondrias perdidas. Similares vestigios se han encontrado en otros flagelados amitocondriales con los estén probablemente emparentados con ellos, por lo que se clasifican juntos en Metamonada. Parabasalia carece de los surcos de alimentación encontrados en los demás grupos, pero esto es probable que también sea una pérdida secundaria.
Los parabasálidos se clasifican en Excavata sobre la base de análisis moleculares, puesto que no presentan ninguno de los caracteres excavados típicos y constituyen un grupo con una morfología única. Hipotéticamente, el antecesor de los parabasálidos era un organismo tetraflagelado, puesto que las cinétidas presentan cuatro cinetosomas privilegiados con apéndices no microtubulares específicos.

Los flagelos están dispuestos en uno o más grupos cerca de la parte anterior de la célula, generalmente en conjuntos de cuatro, aunque en algunos grupos se produce la reducción o pérdida del aparato flagelar, mientras que en otros se produce la multiplicación de todos o parte de los elementos (algunas especies puede llegar a tener hasta mil flagelos). Uno de los flagelos puede adherirse a lo largo de la célula a una proyección de la membrana plasmática formando una membrana ondulante.
Los parabasálidos se caracterizan por presentar un aparato parabasal, constituido por dos o más fibras parabasales estriadas que conectan el aparato de Golgi al sistema flagelar. Esto es, los centrosomas están unidos al prominente aparato de Golgi, distintivo del grupo, mediante las fibras parabasales.
Entre las estructuras de microtúbulos destaca el axostilo, constituido por un conjunto de microtúbulos trenzados que recorre el eje la célula y que en algunos casos se proyectan más allá de esta. Los microtúbulos están dispuestos en una lámina espiralizada que forma un tubo, hueco o relleno, o un cono, a veces rodeando parcialmente el núcleo celular. Sin embargo tiene una estructura diferente de los axostilos de Oxymonadida. Otras estructuras de microtúbulos que pueden presentar los parabasálidos se denominan pelta y costa. La pelta es una estructura de microtúbulos a continuación del axostilo hacia la parte anterior, que recubre parcialmente las estructuras basales de los flagelos. La costa es un haz de microtúbulos dispuesto en el interior de la célula y paralelo a la membrana ondulante que da rigidez a la célula y soporta la membrana.
Los distintos grupos de parabasálidos presentan dos tipos de costa, A y B, que difieren en el patrón de estrías, resultado de la diferente organización de sus subestructuras de fibra. A pesar de sus diferencias, los dos tipos de costa están constituidos por compuestos proteicos similares, por lo que son estructuras homólogas. Otras características que ayudan a distinguir a los distintos grupos son la presencia o ausencia en las cinétidas de una estructura de tipo peine y de un cuerpo infracinetosomial. También se caracterizan en función de la forma del axostilo, que puede ser de tipo Trichomonas o Tritrichomonas.[6]
Clasificación[editar]
Parabasalia se divide en varios grupos que pueden considerarse órdenes o clases.[6][4]
- Los tres primeros órdenes, Trichomonadida, Tritrichomonadida e Hypotrichomonadida tienen grupos de 0-6 flagelos, uno de los cuales parte de un lado de la célula y a menudo forma una membrana ondulada. Usualmente presentan costa. Muchas especies de este orden se encuentran en huéspedes vertebrados, incluyendo Trichomonas vaginalis que causa una enfermedad de transmisión sexual en los seres humanos.
- Los otros órdenes, Cristamonadida, Trichonymphida y Spirotrichonymphida, anteriormente agrupados en Hypermastigida, tienen numerosos grupos de flagelos y se encuentran exclusivamente en el tracto digestivo de insectos. Los flagelos, raramente 4 y más frecuentemente muy numerosos se disponen en un penacho apical o en espirales. La membrana ondulante y la costa están normalmente reducidos o ausentes. Comúnmente presentan múltiples núcleos y axostilos.
Filogenia[editar]
Se ha postulado en 2012 las siguientes relacionesː[7]
| Parabasalia |
| |||||||||||||||||||||||||||
Especies notales[editar]
Algunas especies de interés incluidas en este grupo son las siguientes:
- Trichomonas vaginalis (Trichomonadida), que vive en la vagina de las mujeres.
- Trichomonas tenax, que parasita exclusivamente la cavidad bucal de los seres humanos.
- Trichomonas columbae (Trichomonadida), también llamada T. gallinae, T. diversa, T. halli o T. hepatica, que afecta a aves.[8]
- Pentatrichomonas hominis (Trichomonadida), parásito del intestino grueso de los humanos.
- Tritrichomonas foetus (Tritrichomonadida), también llamada T. bovina, es una enfermedad de transmisión sexual de los bovinos y parásito intestinal en gatos.[9]
- Dientamoeba fragilis (Tritrichomonadida), ameboide parásito de los humanos.
- Histomonas meleagridis (Tritrichomonadida), parásito de las gallinas.
- Mixotricha paradoxa (Cristamonadida), endosimbionte de las termitas.
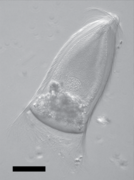
Galería[editar]
-

-

-

-
-
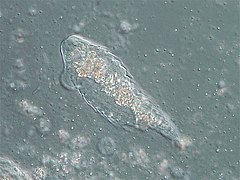 Hypermastigida
Hypermastigida

_Cultured.jpg)



Referencias[editar]
- ↑ Adl, S.M. et al. 2012. The revised classification of eukaryotes. Journal of Eukaryotic Microbiology, 59(5), 429-514
- ↑ Ohkuma M, Iida T, Ohtoko K, et al. (junio de 2005). «Molecular phylogeny of parabasalids inferred from small subunit rRNA sequences, with emphasis on the Hypermastigea». Mol. Phylogenet. Evol. 35 (3): 646-55. PMID 15878133. doi:10.1016/j.ympev.2005.02.013.
- ↑ Ald, S.M. et al. (2007) Diversity, Nomenclature, and Taxonomy of Protists, Syst. Biol. 56(4), 684–689, DOI: 10.1080/10635150701494127.
- ↑ a b Noda, S., Mantini, C., Meloni, D., Inoue, J., Kitade, O., Viscogliosi, E., & Ohkuma, M. (2012). Molecular phylogeny and evolution of parabasalia with improved taxon sampling and new protein markers of actin and elongation factor-1α. PloS one, 7(1), e29938-e29938.
- ↑ Bui, ET; Bradley, PJ; Johnson, PJ (3 de septiembre de 1996). «A common evolutionary origin for mitochondria and hydrogenosomes.». Proceedings of the National Academy of Sciences of the United States of America 93 (18): 9651-6. PMC 38483. PMID 8790385. doi:10.1073/pnas.93.18.9651.
- ↑ a b Gile, G. H., & Slamovits, C. H. (2012). Phylogenetic position of Lophomonas striata Bütschli (Parabasalia) from the hindgut of the cockroach Periplaneta americana. Protist, 163(2), 274-283.
- ↑ Noda S, Mantini C, Meloni D, Inoue J-I, Kitade O, Viscogliosi E, et al. (2012) Molecular Phylogeny and Evolution of Parabasalia with Improved Taxon Sampling and New Protein Markers of Actin and Elongation Factor-1α. PLoS ONE 7(1): e29938. doi:10.1371/journal.pone.0029938
- ↑ Robert M. Stabler. The Journal of Parasitology Vol. 33, No. 3 (Jun., 1947), páginas 207-213, ed. «Trichomonas gallinae, Pathogenic Trichomonad of Birds».
- ↑ Revista de Medicina Veterinaria, Bs As Argentina vol 87: 47-56. 2006. «Tritrichomonas foetus: patogénesis de la mortalidad embrionaria/fetal, caracterización de antígenos vacunales y respuesta inmune inducida». Archivado desde el original el 31 de octubre de 2011.
 Datos: Q134585
Datos: Q134585 Especies: Parabasalia
Especies: Parabasalia