
 French
French Deutsch
Deutschクツコムシ目
| クツコムシ目 | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Cryptocellus goodnighti | |||||||||||||||
| 地質時代 | |||||||||||||||
| 石炭紀(ペンシルベニア紀)–現世 | |||||||||||||||
| 分類 | |||||||||||||||
| |||||||||||||||
| 学名 | |||||||||||||||
| Ricinulei Thorell, 1876 | |||||||||||||||
| 英名 | |||||||||||||||
| Ricinuleid Hooded tickspider | |||||||||||||||
| 下位分類 | |||||||||||||||
| 本文参照 |
クツコムシ目(口籠虫目、学名:Ricinulei) は、鋏角亜門クモガタ綱に属する節足動物の分類群である。クツコムシ(クツコムシ類、ricinuleid)と総称され、節腹類とも言う。小さくずんぐりしたクモに似た姿で、硬い外骨格を持つ。最大の特徴は、前端部に頭蓋(とうがい)と呼ばれる、自動車のボンネットに似た可動式の蓋を持つことである。この蓋は口と鋏角を上から覆うことができる。
分布域は非常に限られており、アフリカ中西部と熱帯アメリカからのみから知られ、2005年までに確認された現生種は全部で1科3属57種と、種類数も大変少ない小さな一群である。和名は、頭蓋を口籠(くつこ:牛馬などが噛み付かないように口にかぶせるカゴ)に見立てたもの。学名「Ricinulei」はラテン語でマダニ類を表す「ricinus」に縮小辞を付けたもので「小さなマダニ」の意という[1]。
形態[編集]

-
 Cryptocellus goodnighti
Cryptocellus goodnighti -
 Cryptocellus sp.
Cryptocellus sp. -
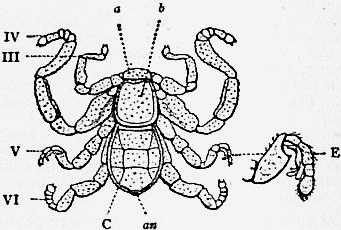 クツコムシの背面(a: 口蓋、b: 背甲、III-VI: 脚、C: 後体、E: オスの第3脚の交接器官)
クツコムシの背面(a: 口蓋、b: 背甲、III-VI: 脚、C: 後体、E: オスの第3脚の交接器官)
.jpg)
_(cropped).jpg)

成体の大きさは5mm~10mmほどの小さな動物である。体は堅く、表面は微細な顆粒に覆われる。前方のやや小さい前体(prosoma)とそれにつながる大きい後体(opisthosoma)の二つの部分からなり、背面から見ると、雪だるま型の胴体に脚を付けたような姿で、全体の形はクモ類に似ている。しかし、背腹に平たく、体の中央がクモ類ほどにはくびれずに密着している。
前体[編集]
前体は頭胸部とも言い、その背面は一枚の背甲に覆われている。背甲の前端部には頭蓋(とうがい、cucullus)またはフード(hood)と呼ばれる可動の板状構造があり、これが蓋のように動いて、上方から口と鋏角を覆い隠すことができる[2]。化石種は多くが背甲の左右に1-2対の側眼を持っていた[3][4]が、現生種はいずれも眼はなく[2]、感覚器としては感覚毛が認められるのみであるが、まったく光を感知できないのか、嗅覚などはどうなのかなどの詳細は未だ不明である。

他のクモガタ類と同様、前体の腹面には鋏角1対、触肢1対、脚4対の計6対の付属肢(関節肢)がある[2]。鋏角は2節の鋏状だが、不動指はやや短く、亜鋏状(牙状、折りたたみナイフ型)に近い形態をもつ[5][6]。触肢は体の下に畳んで左右に張った鎌のようになって、先端に小さな鋏と感覚器がある[7][6][2]。脚は太くがっしりしていて7肢節からなるが、第1脚以外の脚では一部の肢節が複数節に分かれており、跗節(先端の肢節)の場合は第2と第4脚が5節、第3脚が4節で、転節(基部2番目の肢節)の場合は第3と第4脚は2節に分かれる。第2脚は最も長く、感覚に用いられる[2]。脚の先端には2本の爪がある。雄の第3脚は交接器官として複雑な形に変形しており[8]、分類の際の重要な形質となる。前体の腹面は密着した触肢と脚の基節によって占められており、腹板はない[2]。上唇は鋏角と触肢の基節の間に配置され、口はその下の奥に開く[9]。
呼吸器官として気管のみを具えている。クモガタ類の多くでは後体に呼吸器官の開口部があるが、クツコムシでは前体の両後側(第3脚と第4脚の間の上部)に開口していて[10][2]、そのすぐ内側で無数の細い気管に分かれている。気門の位置は直後の後体の左右に覆われるため、後体を除去しない限り外から観察できない[11]。排出はマルピーギ管と第3脚基部後方にある脚基腺(coxal gland)による。神経系は各神経節の境目は不明瞭で、前体に融合した中枢神経系になっている[9]。
後体[編集]
_(cropped).jpg)

後体はすなわち腹部で、付属肢らしい構造はない[2]。背面と腹面の外骨格(背板と腹板)はそれぞれ8節と9節に見えるが、発生学的証拠がない上で、その一部も明らかに複数の体節の癒合や変形が進んだ結果であるため、正確の体節数を明らかにするのが難しい[2]。
第1節の背板は前後で幅狭く、背甲に隠れて見立てない。前半部の凹凸は背甲の後縁と相互に嵌め込まれ、背甲としっかりと密着している「locking ridge」となる[5][2]。
見かけ上の次の「4節」は、それぞれ(少なくとも後方の大きな3節が[2])前後2対の内突起(体節ごとに1対のみをもつ筋肉の接続点)をもつことから、2つの体節から癒合していた重体節(diplosegment)と考えられる[5][2]。これらの4つの重体節の背板は横長く、それぞれ中央の大きい板と左右の小さい板とに3分されているため、背面全体では大小12枚の背板からなるように見える[5]。腹板は背板ほど明瞭には小分割されず、比較的単純な形で重体節ごとに並んでいる[5]。化石種では、これらの全ての節が1つの甲羅のように癒合した場合がある[12]。

腹面の見かけ上最初2つの腹板は非常に小さく、ひし形の構造体にまとまり、その間には横長の生殖口がある[13]が、これはクモガタ類の生殖口が、知る限り必ず第2節上にある[2]のとは異なっている。このことから、クツコムシ類の生殖口のすぐ前にある見かけ上の「第1腹板」は実は第2節の腹板、もしくはそれが外見通り第1節由来で、第2節の腹板が部分的に退化消失し、第2節の生殖口を第1腹板に隣接させたと考えられる[2]。
最終の3節は円筒状で、望遠鏡のように伸縮可能[13][9][2]な小さいイボ状を構成し、文献によっては尾部(pygidium)、後腹部(postabdomen)、終体(metasoma)[9]などと呼ばれる[2]。後端に肛門があり、尾節はない[2]。
上述の解釈をまとめると、クツコムシ類の後体は発生学的に12節(第1節+2節を含んだ重体節4つ+最終3節)からなると推測される[2]。
| 後体の体節 | 1 | 2+3 | 4+5 | 6+7 | 8+9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|
| 構造 | 背板 腹板? | 背板(横3枚) 第2節由来の腹板? 生殖口 第3節由来の腹板 | 背板(横3枚) 腹板 | 背板(横3枚) 腹板 | 背板(横3枚) 腹板 | 円筒状 | 円筒状 | 円筒状 |
生態[編集]
ほとんどの種は熱帯雨林内の落ち葉の下などに生息するが、洞窟に棲むものもいくつか知られている。いずれも適度の湿度が保たれていることが重要らしい。動作は遅く、第2脚を触角のようにして探りながら歩く。捕食性で、主に他の小さな節足動物などを食べているとされる。配偶行動については詳しくは知られていないが、雄が交接用の第3脚を使って精包を雌に渡すのが観察されている。雌は卵を頭蓋の下に保護して持ち運ぶ[8]。生まれたばかりの幼生はダニ類のそれと同様に3対の歩脚をもち、後に脱皮して親と同じ4対の歩脚となる。
生息域が限られるため、かつては非常に稀な動物と考えられていたが、その後の調査研究の結果、その生息地では極めて普通に見られる場合が多いことがわかった。しかし分布域の一部、特にアフリカの生息域は、森林の伐採などによってその生息が脅かされている可能性も指摘されている。古生代石炭紀(主にペンシルベニア紀)から化石が出ており、当時の個体数は多かったらしく、化石として発見されている個体数と現生で確認された個体数とに大差がないと言われていた時期もある。
分類[編集]
多くのクモガタ類と同様、クツコムシ類の系統的位置は明確でない[2]。現生群に限れば、形態学的には同様に3対の脚をもつ幼生の形態に基づいて、ダニ類の近縁群と位置付けられていた[14][2]。クモガタ類の中で最も基盤的な存在と見られることもあったが、頭蓋その他の派生的な形質が多いことから、むしろ系統的に新しい群であると考えられるようになった[要出典]。分子系統学的見解でもその位置を明らかにせず、ヒヨケムシ類に近縁[15][16][17]、ザトウムシ類に近縁[18]、さらにクモガタ類でないカブトガニ類に近縁[19][20]など、様々な解析結果が出ている。
.jpg)
.jpg)

また、3つの外骨格に分かれた後体の背板・先端にはさみを持った触肢・凹凸に嵌め込まれた背甲と第1背板の繋ぎ目などの形質は、絶滅したクモガタ類であるワレイタムシ類と共通しており、類縁関係にあることが示唆される[5][6][21]。ワレイタムシ類自体は四肺類(クモ・ウデムシ・サソリモドキなどを含んだ系統群)の近縁として広く認められるため、もしクツコムシ類とワレイタムシ類は近縁であれば、クツコムシ類自体も四肺類の近縁と見なされる[5][6]。この系統仮説に踏まえて、クツコムシ類の亜鋏状に近いはさみ型の鋏角は、クモガタ類として祖先的な鋏状の鋏角と四肺類の折りたたみナイフ型の鋏角の中間形質を表した可能性も挙げられる[5][6]。
下位分類[編集]
.jpg)
a:Ricinoides atewa の幼生
b:Ricinoides karschii の雄
c:Pseudocellus pearsei の雄
d:Cryptocellus becki の雌と2匹の幼生
e:Cryptocellus 属の未記載種の雌
.jpg)
Selden (1992)は化石種と現生種とをそれぞれ別亜目として分けて2亜目を創設したが、この分類によれば現生種はシンクツコムシ亜目のクツコムシ科のみからなる。さらにクツコムシ科はかつては西アフリカ産のRicinoides属と中南米産のCryptocellusの2属に分けられていたが、 Platnick (1980)が中米産の一部をPseudocellusという新属として分けたことで3属とされるようになった。一方、化石種はムカシクツコムシ亜目に属する2科からなり、長らく2属とされてきたが、Selden(1992)が新たに2属を創設して4属となっている。しかし研究者自体が少なく、分布域が限られることや、小型であることなどもあって生態学的にも分類学的にも研究は十分ではなく、今後の研究の進展によって種数や分類の内容、他のクモガタ類との系統関係、生態学的知見などが大幅に追加修正される可能性もある。
以下は2005年時点までに知られている分類の概要である。
クツコムシ目 Ricinulei (現生種:3属57種・化石種:4属16種)
- シンクツコムシ亜目 Neoricinulei Selden 1992 (全て現生)
- †ムカシクツコムシ亜目 Palaeoricinulei Selden 1992 (全て化石種)
- (上科)Curculioidoidea
- (科)Curculioididae Cockerell, 1916 (2属11種)
- (属)Amarixys Selden, 1992 (3種)
- (属)Curculioides Buckland, 1837 (8種)
- (科)Poliocheridae Scudder, 1884 (2属5種)
- (属)Poliochera Scudder, 1884 (4種)
- (属)Terpsicroton Selden, 1992 (1種)
- (科)Curculioididae Cockerell, 1916 (2属11種)
- (上科)Curculioidoidea
脚注[編集]
- ^ 『日本産クモ類』p.12
- ^ a b c d e f g h i j k l m n o p q r s t u A., Dunlop, Jason; C., Lamsdell, James (2017). “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3). ISSN 1467-8039.
- ^ Seiden, Paul A. (1992/ed). “Revision of the fossil ricinuleids” (英語). Earth and Environmental Science Transactions of The Royal Society of Edinburgh 83 (4): 595–634. doi:10.1017/S0263593300003333. ISSN 1473-7116.
- ^ Miether, Sebastian T.; Dunlop, Jason A. (2016/07). “Lateral eye evolution in the arachnids”. Arachnology 17 (2): 103–119. doi:10.13156/arac.2006.17.2.103. ISSN 2050-9928.
- ^ a b c d e f g h Dunlop, J. A. (1996) (English). Evidence for a sister group relationship between Ricinulei and Trigonotarbida
- ^ a b c d e Dunlop, Jason A. (2009年). “A fossil trigonotarbid arachnid with a ricinuleid-like pedipalpal claw” (英語). undefined. 2018年11月29日閲覧。
- ^ Talarico, Giovanni (2008年). “The pedipalp of Pseudocellus pearsei (Ricinulei, Arachnida) - ultrastructure of a multifunctional organ.” (英語). undefined. 2018年11月29日閲覧。
- ^ a b McLean, Callum J.; Garwood, Russell J.; Brassey, Charlotte A. (2018-11-06). “Sexual dimorphism in the Arachnid orders” (英語). PeerJ 6: e5751. doi:10.7717/peerj.5751. ISSN 2167-8359. PMC 6225839. PMID 30416880.
- ^ a b c d Talarico, Giovanni; Lipke, Elisabeth; Alberti, Gerd (2010). “Gross morphology, histology, and ultrastructure of the alimentary system of Ricinulei (Arachnida) with emphasis on functional and phylogenetic implications” (英語). Journal of Morphology 272 (1): 89–117. doi:10.1002/jmor.10897. ISSN 1097-4687.
- ^ Ax, Peter (2000). Ax, Peter. ed (英語). Multicellular Animals: The Phylogenetic System of the Metazoa. Volume II. Berlin, Heidelberg: Springer. pp. 130–135. doi:10.1007/978-3-662-10396-8_36. ISBN 978-3-662-10396-8
- ^ Adis, Joachim; Messner, Benjamin; Platnick, Norman (1999-04-01). “Morphological Structures and Vertical Distribution in the Soil Indicate Facultative Plastron Respiration in Cryptocellus adisi (Arachnida, Ricinulei) from Central Amazonia”. Studies on Neotropical Fauna and Environment 34 (1): 1–9. doi:10.1076/snfe.34.1.1.8915. ISSN 0165-0521.
- ^ Pocock, R. I. (1911). A monograph of the terrestrial Carboniferous Arachnida of Great Britain. London :: Printed for the Palæontographical Society,
- ^ a b Talarico, G.; García Hernández, L. F.; Michalik, P. (2008-09-01). “The male genital system of the New World Ricinulei (Arachnida): Ultrastructure of spermatozoa and spermiogenesis with special emphasis on its phylogenetic implications” (英語). Arthropod Structure & Development 37 (5): 396–409. doi:10.1016/j.asd.2008.01.006. ISSN 1467-8039.
- ^ Shultz, Jeffrey W. (2007). A phylogenetic analysis of the arachnid orders based on morphological characters. doi:10.1111/J.1096-3642.2007.00284.X.
- ^ Regier, Jerome C.; Shultz, Jeffrey W.; Zwick, Andreas; Hussey, April; Ball, Bernard; Wetzer, Regina; Martin, Joel W.; Cunningham, Clifford W. (2010-02). “Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences” (英語). Nature 463 (7284): 1079–1083. doi:10.1038/nature08742. ISSN 1476-4687.
- ^ Sharma, Prashant P.; Kaluziak, Stefan T.; Pérez-Porro, Alicia R.; González, Vanessa L.; Hormiga, Gustavo; Wheeler, Ward C.; Giribet, Gonzalo (2014-08-08). “Phylogenomic Interrogation of Arachnida Reveals Systemic Conflicts in Phylogenetic Signal” (英語). Molecular Biology and Evolution 31 (11): 2963–2984. doi:10.1093/molbev/msu235. ISSN 1537-1719.
- ^ Howard, Richard J.; Puttick, Mark N.; Edgecombe, Gregory D.; Lozano-Fernandez, Jesus (2020-11-01). “Arachnid monophyly: Morphological, palaeontological and molecular support for a single terrestrialization within Chelicerata” (英語). Arthropod Structure & Development 59: 100997. doi:10.1016/j.asd.2020.100997. ISSN 1467-8039.
- ^ Lozano-Fernandez, Jesus; Tanner, Alastair R.; Giacomelli, Mattia; Carton, Robert; Vinther, Jakob; Edgecombe, Gregory D.; Pisani, Davide (2019-05-24). “Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida” (英語). Nature Communications 10 (1): 2295. doi:10.1038/s41467-019-10244-7. ISSN 2041-1723.
- ^ Sharma, Prashant P.; Kaluziak, Stefan T.; Pérez-Porro, Alicia R.; González, Vanessa L.; Hormiga, Gustavo; Wheeler, Ward C.; Giribet, Gonzalo (2014-11-01). “Phylogenomic Interrogation of Arachnida Reveals Systemic Conflicts in Phylogenetic Signal” (英語). Molecular Biology and Evolution 31 (11): 2963–2984. doi:10.1093/molbev/msu235. ISSN 0737-4038.
- ^ Ballesteros, Jesús A.; Sharma, Prashant P. (2019). “A Critical Appraisal of the Placement of Xiphosura (Chelicerata) with Account of Known Sources of Phylogenetic Error” (英語). Systematic Biology. doi:10.1093/sysbio/syz011.
- ^ Dunlop, J. A.; Alberti, G. (2007-09-04). “The affinities of mites and ticks: a review” (ドイツ語). Journal of Zoological Systematics and Evolutionary Research 0 (0): 070907105857006–???. doi:10.1111/j.1439-0469.2007.00429.x. ISSN 0947-5745.
参考文献[編集]
- Harvey,Mark S., 2002. The neglected counsis:What do we know about the smaller arachnid orders? The Journal of Arachnology 30: 357-372. PDF
- Selden,P.A., 1992. Revision of the fossil ricinuleids. Transactions of the Royal Society of Edinburgh,Earth Sciences 83: 595-634.
- 内田亨『動物系統分類学』 第7巻 中 A、中山書店、1966年。ISBN 4-521-07111-2。
- 小野展嗣『日本産クモ類』東海大学出版会、2009年。ISBN 978-4-486-01791-2。
関連項目[編集]
外部リンク[編集]
- Photos of Ricinulei - クツコムシの画像
- Morphology & Evolution of Arachnids - クツコムシ電子顕微鏡画像(2007年3月13日時点のアーカイブ)
- SYNOPSIS OF THE DESCRIBED RICINULEI OF THE WORLD - クツコムシ目リスト